Enzimas
Conformación en
forma de diagrama de cintas rodeado por el modelo
de relleno de espacio de la proteína. Esta proteína
es una eficiente enzima involucrada en el proceso de transformación de azúcares
en energía
en las células.
Las enzimas son moléculas
de naturaleza proteica que catalizan
reacciones químicas, siempre que sea termodinámicamente
posible (si bien no pueden hacer que el proceso sea más termodinámicamente
favorable).En estas reacciones, las enzimas actúan sobre unas moléculas
denominadas sustratos, las cuales se convierten en
moléculas diferentes denominadas productos. Casi todos los procesos en las células
necesitan enzimas para que ocurran a unas tasas significativas. A las
reacciones mediadas por enzimas se las denomina reacciones
enzimáticas.
Debido a que
las enzimas son extremadamente selectivas con sus sustratos y su velocidad
crece sólo con algunas reacciones, el conjunto (set) de enzimas
sintetizadas en una célula determina el tipo de metabolismo
que tendrá cada célula. A su vez, esta síntesis depende de la regulación de la
expresión génica.
Como todos los catalizadores,
las enzimas funcionan disminuyendo la energía de activación (ΔG‡) de
una reacción, de forma que se acelera sustancialmente la tasa de reacción. Las
enzimas no alteran el balance energético de las reacciones en que intervienen,
ni modifican, por lo tanto, el equilibrio de la reacción, pero consiguen
acelerar el proceso incluso millones de veces. Una reacción que se produce bajo
el control de una enzima, o de un catalizador en general, alcanza el equilibrio
mucho más deprisa que la correspondiente reacción no catalizada.
Al igual que
ocurre con otros catalizadores, las enzimas no son consumidas por las
reacciones que catalizan, ni alteran su equilibrio químico. Sin embargo, las
enzimas difieren de otros catalizadores por ser más específicas. Las enzimas
catalizan alrededor de 4.000 reacciones bioquímicas distintas. No todos los
catalizadores bioquímicos son proteínas, pues algunas moléculas de ARN son capaces de
catalizar reacciones (como la subunidad 16S de los ribosomas
en la que reside la actividad peptidil transferasa).También cabe
nombrar unas moléculas sintéticas denominadas enzimas artificiales
capaces de catalizar reacciones químicas como las enzimas clásicas.
La actividad de
las enzimas puede ser afectada por otras moléculas. Los inhibidores enzimáticos son moléculas que
disminuyen o impiden la actividad de las enzimas, mientras que los activadores
son moléculas que incrementan dicha actividad. Asimismo, gran cantidad de
enzimas requieren de cofactores para su actividad. Muchas drogas o fármacos son
moléculas inhibidoras. Igualmente, la actividad es afectada por la temperatura,
el pH, la concentración
de la propia enzima y del sustrato, y otros factores físico-químicos.
Algunas enzimas
son usadas comercialmente, por ejemplo, en la síntesis de antibióticos
y productos domésticos de limpieza. Además, son ampliamente utilizadas en
diversos procesos industriales, como son la fabricación de alimentos,
destinción de jeans
o producción de biocombustibles.
Estructuras y mecanismos
Diagrama de
cintas que representa la estructura de una anhidrasa carbónica de tipo II. La esfera gris
representa al cofactor
zinc situado en el centro
activo. Las enzimas son
generalmente proteínas globulares que pueden presentar tamaños muy variables,
desde 62 aminoácidos como en el caso del monómero
de la 4-oxalocrotonato
tautomerasa,15 hasta los 2.500
presentes en la sintasa de ácidos grasos.
Las actividades
de las enzimas vienen determinadas por su estructura tridimensional, la cual
viene a su vez determinada por la secuencia de aminoácidos.Sin embargo, aunque
la estructura determina la función, predecir una nueva actividad enzimática
basándose únicamente en la estructura de una proteína es muy difícil, y un
problema aún no resuelto.Casi todas las
enzimas son mucho más grandes que los sustratos sobre los que actúan, y solo
una pequeña parte de la enzima (alrededor de 3 a 4 aminoácidos)
está directamente involucrada en la catálisis. La región que
contiene estos residuos encargados de catalizar la reacción es denominada centro
activo.
Las enzimas también pueden contener sitios con la capacidad de unir
cofactores,
necesarios a veces en el proceso de catálisis,
o de unir pequeñas moléculas, como los sustratos o productos (directos o
indirectos) de la reacción catalizada. Estas uniones de la enzima con sus
propios sustratos o productos pueden incrementar o disminuir la actividad
enzimática, dando lugar así a una regulación por retroalimentación positiva o negativa, según el
caso. Al igual que
las demás proteínas, las enzimas se componen de una cadena lineal de aminoácidos
que se pliegan durante el proceso de traducción para dar lugar a una estructura terciaria tridimensional de la
enzima, susceptible de presentar actividad. Cada secuencia de aminoácidos es
única y por tanto da lugar a una estructura única, con propiedades únicas. En
ocasiones, proteínas individuales pueden unirse a otras proteínas para formar
complejos, en lo que se denomina estructura cuaternaria de las proteínas.
La mayoría de
las enzimas, al igual que el resto de las proteínas, pueden ser
desnaturalizadas si se ven sometidas a agentes desnaturalizantes como el calor, los pHs extremos o ciertos
compuestos como el SDS.
Estos agentes destruyen la estructura terciaria de las proteínas de forma
reversible o irreversible, dependiendo de la enzima y de la condición.
Especificidad
Las enzimas
suelen ser muy específicas tanto del tipo de reacción que catalizan como del sustrato
involucrado en la reacción. La forma, la carga y las características hidrofílicas/hidrofóbicas
de las enzimas y los sustratos son los responsables de dicha especificidad. Las
enzimas también pueden mostrar un elevado grado de estereoespecificidad, regioselectividad
y quimioselectividad.
Algunas
de estas enzimas que muestran una elevada especificidad y precisión en su
actividad son aquellas involucrados en la replicación
y expresión
del genoma.
Estas enzimas tienen eficientes sistemas de comprobación y corrección de
errores, como en el caso de la ADN
polimerasa, que cataliza una reacción de replicación en un primer
paso, para comprobar posteriormente si el producto obtenido es el correcto.21 Este proceso, que
tiene lugar en dos pasos, da como resultado una media de tasa de error
increíblemente baja, en torno a 1 error cada 100 millones de reacciones en
determinadas polimerasas de mamíferos.Este tipo de
mecanismos de comprobación también han sido observados en la ARN
polimerasa,en la ARNt
aminoacil sintetasa y en la actividad
de selección de los aminoacil-tRNAs.




Gráfica de las energías de las diferentes fases de una reacción química. Los sustratos precisan mucha energía para alcanzar el estado de transición, pero una vez alcanzado, se transforman en productos. La enzima estabiliza el estado de transición, reduciendo la energía necesaria para formar los productos.







Un ejemplo de esto es el tipo más común de fenilcetonuria. En esta enfermedad genética se produce una mutación de un único aminoácido en la fenilalanina hidroxilasa, una enzima que cataliza la primera reacción de la ruta de degradación de la fenilalanina y de compuestos relacionados. Al ser esta enzima inactiva, se acumulan una serie de productos que terminan dando lugar a la aparición de retardo mental si no se recibe tratamiento.
Aquellas
enzimas que producen metabolitos secundarios son denominadas
promiscuas, ya que pueden actuar sobre una gran variedad de sustratos. Por
ello, se ha sugerido que esta amplia especificidad de sustrato podría ser clave
en la evolución y diseño de nuevas rutas biosintéticas.
Modelo de la "llave-cerradura"
Las enzimas son
muy específicas, como sugirió Emil Fisher en 1894. Con base a sus
resultados dedujo que ambas moléculas, enzima y sustrato, poseen
complementariedad geométrica, es decir, sus estructuras encajan exactamente una
en la otra,por lo que ha sido denominado como modelo de la "llave-cerradura",
refiriéndose a la enzima como a una especie de cerradura y al sustrato como a
una llave que encaja de forma perfecta en dicha cerradura. Sin embargo, si bien
este modelo explica la especificidad de las enzimas, falla al intentar explicar
la estabilización del estado de transición que logran adquirir
las enzimas.
Modelo del encaje inducido
Diagrama que
esquematiza el modo de acción del modelo del encaje inducido.
En 1958 Daniel Koshland sugiere
una modificación al modelo de la llave-cerradura: las enzimas son estructuras
bastante flexibles y así el sitio activo podría cambiar su conformación
estructural por la interacción con el sustrato. Como resultado de
ello, la cadena
aminoacídica que compone el sitio activo es moldeada en posiciones
precisas, lo que permite a la enzima llevar a cabo su función catalítica. En
algunos casos, como en las glicosidasas, el sustrato
cambia ligeramente de forma para entrar en el sitio activo. El sitio activo
continua dicho cambio hasta que el sustrato está completamente unido, momento
en el cual queda determinada la forma y la carga final.
Mecanismos
Las enzimas
pueden actuar de diversas formas, aunque, como se verá a continuación, siempre
dando lugar a una disminución del valor de ΔG‡:
·
Reducción de la
energía de activación mediante la creación
de un ambiente en el cual el estado de transición es estabilizado (por ejemplo,
forzando la forma de un sustrato: la enzima produce un cambio de conformación
del sustrato unido el cual pasa a un estado de transición, de modo que ve
reducida la cantidad de energía que precisa para completar la transición).
·
Reduciendo la
energía del estado de transición, sin afectar la forma del sustrato, mediante
la creación de un ambiente con una distribución de carga óptima para que se
genere dicho estado de transición.
·
Proporcionando
una ruta alternativa. Por ejemplo, reaccionando temporalmente con el sustrato
para formar un complejo intermedio enzima/sustrato (ES), que no sería factible
en ausencia de enzima.
·
Reduciendo la
variación de entropía de la reacción mediante la acción de orientar
correctamente los sustratos, favoreciendo así que se produzca dicha reacción.
·
Incrementando
la velocidad de la enzima mediante un aumento de temperatura. El incremento de
temperatura facilita la acción de la enzima y permite que se incremente su
velocidad de reacción. Sin embargo, si la temperatura se eleva demasiado, la
conformación estructural de la enzima puede verse afectada, reduciendo así su
velocidad de reacción, y sólo recuperando su actividad óptima cuando la
temperatura se reduce. No obstante, algunas enzimas son termolábiles y trabajan
mejor a bajas temperaturas.Cabe destacar
que este efecto entrópico implica la desestabilización del estado basal, y su contribución a
la catálisis es relativamente pequeña.
Estabilización del estado de transición
La comprensión
del origen de la reducción del valor de ΔG‡ en una reacción
enzimática requiere elucidar previamente cómo las enzimas pueden estabilizar su
estado de transición, más que el estado de transición de la reacción.
Aparentemente, la forma más efectiva para alcanzar la estabilización es la
utilización de fuerzas electrostáticas,
concretamente, poseyendo un ambiente polar relativamente fijado que pueda
orientarse hacia la distribución de carga del estado de transición. Ese tipo de
ambientes no existen ni se generan en ausencia de enzimas.
Dinámica y funció
La dinámica interna
se define como el movimiento de diferentes partes de la estructura de la
enzima, desde residuos individuales de aminoácidos,
hasta grupos de aminoácidos o incluso un dominio
proteico entero. Estos movimientos se producen a diferentes escalas
de tiempo que van desde femtosegundos hasta segundos.
Casi cualquier residuo de la estructura de la enzima puede contribuir en el
proceso de catálisis por medio de movimientos dinámicos.
Los movimientos de
las proteínas son vitales en muchas enzimas. Dichos movimientos podrán ser más
o menos importantes según si los cambios conformacionales se producen por
vibraciones pequeñas y rápidas o grandes y lentas, y dicha importancia
dependerá del tipo de reacción que lleve a cabo la enzima. Sin embargo, aunque
estos movimientos son importantes en el proceso de unión y liberación de
sustratos y productos, aún no está claro si estos movimientos ayudan a acelerar
los pasos químicos de las reacciones enzimáticas.Estos nuevos
avances también tienen implicaciones en la comprensión de los efectos
alostéricos y en el desarrollo de nuevos fármacos.
Modulación alostérica

Transición
alostérica de una enzima entre los estados R y T, estabilizada por un agonista,
un inhibidor y un sustrato.
Los sitios alostéricos
son zonas de la enzima con capacidad de reconocer y unir determinadas moléculas
en la célula. Las uniones a las que dan lugar son débiles y no covalentes,
y generan un cambio en la conformación estructural de la enzima que repercute
en el sitio activo, afectando así a la velocidad de reacción de la enzima.Las interacciones alostéricas pueden tanto
inhibir como activar enzimas, y son una forma muy común de controlar las
enzimas en las células.
Cofactores y coenzimas
Cofactores
Algunas enzimas
no precisan ningún componente adicional para mostrar una total actividad. Sin
embargo, otras enzimas requieren la unión de moléculas no proteicas denominadas
cofactores
para poder ejercer su actividad.Los cofactores
pueden ser compuestos inorgánicos, como los iones metálicos y los complejos
ferrosulfurosos, o compuestos orgánicos, como la flavina
o el grupo hemo.
Los cofactores orgánicos pueden ser a su vez grupos prostéticos, que se unen fuertemente a
la enzima, o coenzimas,
que son liberados del sitio activo de la enzima durante la reacción. Las
coenzimas incluyen compuestos como el NADH, el NADPH y el adenosín trifosfato. Estas moléculas
transfieren grupos funcionales entre enzimas.
Un ejemplo de
una enzima que contiene un cofactor es la anhidrasa carbónica, en la cual el zinc (cofactor) se
mantiene unido al sitio activo, tal y como se muestra en la figura anterior (situada al inicio de la sección "Estructuras y mecanismos").Estas moléculas
suelen encontrarse unidas al sitio activo y están implicadas en la catálisis.
Por ejemplo, la flavina y el grupo hemo suelen estar implicados en reacciones redox.
Las enzimas que
requieren un cofactor pero no lo tienen unido son denominadas apoenzimas
o apoproteínas. Una apoenzima junto con cofactor(es) es denominada holoenzima
(que es la forma activa). La mayoría de los cofactores no se unen covalentemente
a sus enzimas, pero sí lo hacen fuertemente. Sin embargo, los grupos
prostéticos pueden estar covalentemente unidos, como en el caso de la tiamina pirofosfato en la
enzima piruvato deshidrogenasa. El término
"holoenzima" también puede ser aplicado a aquellas enzimas que
contienen múltiples subunidades, como en el caso de la ADN
polimerasa, donde la holoenzima es el complejo con todas las
subunidades necesarias para llevar a cabo la actividad enzimática.
Coenzimas

{kind=link}
Modelo
tridimensional de esferas de la coenzima NADH.
Las coenzimas
son pequeñas moléculas orgánicas que transportan grupos químicos de una enzima
a otra.Algunos de estos compuestos, como la riboflavina,
la tiamina
y el ácido fólico son vitaminas
(las cuales no pueden ser sintetizados en cantidad suficiente por el cuerpo
humano y deben ser incorporados en la dieta). Los grupos químicos
intercambiados incluyen el ion hidruro (H-) transportado por NAD o NADP+, el
grupo fosfato
transportado por el ATP, el grupo acetilo
transportado por la coenzima A, los grupos formil, metenil o metil
transportados por el ácido fólico
y el grupo metil transportado por la S-Adenosil metionina.
Debido a que
las coenzimas sufren una modificación química como consecuencia de la actividad
enzimática, es útil considerar a las coenzimas como una clase especial de
sustratos, o como segundos sustratos, que son comunes a muchas enzimas
diferentes. Por ejemplo, se conocen alrededor de 700 enzimas que utilizan la
coenzima NADH
Las coenzimas
suelen estar continuamente regenerándose y sus concentraciones suelen
mantenerse a unos niveles fijos en el interior de la célula: por ejemplo, el
NADPH es regenerado a través de la ruta de las pentosas fosfato y la S-Adenosil
metionina por medio de la metionina
adenosiltransferasa. Esta regeneración continua significa que
incluso pequeñas cantidades de coenzimas son utilizadas intensivamente. Por
ejemplo, el cuerpo humano gasta su propio peso en ATP cada día.
Termodinámica

Gráfica de las energías de las diferentes fases de una reacción química. Los sustratos precisan mucha energía para alcanzar el estado de transición, pero una vez alcanzado, se transforman en productos. La enzima estabiliza el estado de transición, reduciendo la energía necesaria para formar los productos.
Al igual que
sucede con todos los catalizadores, las enzimas no alteran el
equilibrio químico de la reacción. Generalmente, en presencia de una enzima, la
reacción avanza en la misma dirección en la que lo haría en ausencia de enzima,
sólo que más rápido. Sin embargo, en ausencia de enzima, podría producirse una
reacción espontánea que generase un producto diferente debido a que en esas
condiciones, dicho producto diferente se forma más rápidamente.
Además, las
enzimas pueden acoplar dos o más reacciones, por lo que una reacción
termodinámicamente favorable puede ser utilizada para favorecer otra reacción
termodinámicamente desfavorable. Por ejemplo, la hidrólisis
de ATP suele ser utilizada para favorecer otras
reacciones químicas
Las enzimas
catalizan reacciones químicas tanto en un sentido como en el contrario. Nunca
alteran el equilibrio, sino únicamente la velocidad a la que es alcanzado. Por
ejemplo, la anhidrasa carbónica cataliza su reacción en una
u otra dirección dependiendo de la concentración de los reactantes, como se
puede ver a continuación:

(en tejidos; alta concentración de CO2)

(en pulmones;
baja concentración de CO2)
Si el
equilibrio se ve muy desplazado en un sentido de la reacción, es decir, se
convierte en una reacción muy exergónica, la reacción se hace
efectivamente irreversible. Bajo estas condiciones, la enzima únicamente
catalizará la reacción en la dirección permitida desde un punto de vista
termodinámico.
Cinética
Artículo principal: Cinética enzimática

{kind=link}
Mecanismo para
una reacción catalizada por una enzima con un único sustrato. La enzima (E) une
un sustrato (S) y genera un producto (P).
La cinética enzimática es el estudio de cómo las
enzimas se unen a sus sustratos y los transforman en productos. Los datos de
equilibrios utilizados en los estudios cinéticos son obtenidos mediante ensayos enzimáticos.
En 1902, Victor Henri propuso una teoría
cuantitativa sobre la cinética enzimática, pero sus datos experimentales no
fueron muy útiles debido a que la importancia de la concentración del ion de hidrógeno
aún no era considerada. Después de que Peter Lauritz Sørensen definiera la escala
logarítmica del pH
e introdujera el concepto de "tampón" (buffer) en 1909, el químico alemán Leonor
Michaelis y su postdoctoral canadiense Maud Leonora Menten repitieron los experimentos
de Henri confirmando su ecuación, que actualmente es conocida como cinética de
Henri-Michaelis-Menten (o simplemente cinética de Michaelis-Menten). Su trabajo fue
desarrollado más en profundidad por George Edward Briggs y J. B. S.
Haldane, quienes obtuvieron las ecuaciones cinéticas que se
encuentran tan ampliamente extendidas en la actualidad.
La mayor
contribución de Henri fue la idea de dividir las reacciones enzimáticas en dos
etapas. En la primera, el sustrato se une reversiblemente a la enzima, formando
el complejo enzima-sustrato (también denominado complejo Michaelis). En la
segunda, la enzima cataliza la reacción y libera el producto.

Curva de
saturación de una reacción enzimática donde se muestra la relación entre la
concentración de sustrato y la velocidad de la reacción.
Las enzimas
pueden catalizar hasta varios millones de reacciones por segundo. Por ejemplo,
la descarboxilación no enzimática de la orotidina 5'-monofosfato
tiene una vida media de 78 millones de años. Sin embargo, cuando la enzima orotidina
5'-fosfato descarboxilasa está presente en el medio, ese mismo
proceso tarda apenas 25 milisegundos.
Las velocidades de
las enzimas dependen de las condiciones de la solución y de la concentración de
sustrato. Aquellas condiciones que desnaturalizan una proteína, como
temperaturas elevadas, pHs extremos o altas concentraciones de sal, dificultan
o impiden la actividad enzimática, mientras que elevadas concentraciones de
sustrato tienden a incrementar la actividad. Para encontrar la máxima velocidad
de una reacción enzimática, la concentración de sustrato se incrementa hasta
que se obtiene una tasa constante de formación de producto (véase la curva de
saturación representada en la figura de la derecha). La saturación ocurre
porque, cuando la concentración de sustrato aumenta, disminuye la concentración
de enzima libre, que se convierte en la forma con sustrato unido (ES). A la
máxima velocidad (Vmax) de la enzima, todos los sitios
activos de dicha enzima tienen sustrato unido, y la cantidad de complejos ES es
igual a la cantidad total de enzima. Sin embargo, Vmax es
sólo una de las constantes cinéticas de la enzima. La cantidad de sustrato
necesario para obtener una determinada velocidad de reacción también es
importante.
Este parámetro viene dado por la constante de Michaelis-Menten (Km),
que viene a ser la concentración de sustrato necesaria para que una enzima
alcance la mitad de su velocidad máxima. Cada enzima tiene un valor de Km
característico para un determinado sustrato, el cual puede decirnos cómo de
afín es la unión entre el sustrato y la enzima. Otra constante útil es kcat,
que es el número de moléculas de sustrato procesadas por cada sitio activo por
segundo.
La eficiencia
de una enzima puede ser expresada en términos de kcat/Km,
en lo que se denomina constante de especificidad, que incorpora la constante de velocidad de todas las fases
de la reacción. Debido a que la constante de especificidad contempla tanto la
afinidad como la capacidad catalítica, es un parámetro muy útil para comparar
diferentes enzimas o la misma enzima con diferentes sustratos. El valor máximo
teórico de la constante de especificidad es denominado límite de difusión tiene
un valor de 108-109 (M-1 s-1).
Llegados a este punto, cada colisión de la enzima con su sustrato da lugar a la
catálisis, con lo que la velocidad de formación de producto no se ve limitada
por la velocidad de reacción, sino por la velocidad de difusión. Las enzimas
que poseen esta propiedad son llamadas enzimas
catalíticamente perfectas o cinéticamente perfectas.
Ejemplos de este tipo de enzimas son la triosa fosfato isomerasa, la anhidrasa carbónica, la acetilcolinesterasa, la catalasa,
la fumarasa, la beta-lactamasa
y la superóxido dismutasa.
La cinética de
Michaelis-Menten depende de la ley de acción de masas, que se deriva
partiendo de los supuestos de difusión
libre y colisión al azar. Sin embargo, muchos procesos bioquímicos o celulares
se desvían significativamente de estas condiciones, a causa de fenómenos como
el crowding macromolecular,
la separación de etapas entre enzima-sustrato-producto, o los movimientos
moleculares uni- o bidimensionales. No obstante, en
estas situaciones se puede aplicar una cinética de Michaelis-Menten fractal.
Algunas enzimas
presentan una cinética más rápida que la velocidad de difusión, lo que en
principio parecería ser imposible. Se han propuesto diversos mecanismos para
tratar de explicar este fenómeno. Uno de los modelos propone que algunas
proteínas podrían tener la capacidad de acelerar la catálisis secuestrando el
sustrato y orientándolo mediante campos eléctricos dipolares. Otro modelo
propone un mecanismo de efecto túnel cuántico, donde un protón o un
electrón pueden formar un túnel a través de barreras de activación, aunque
existe cierta controversia en cuanto al efecto túnel que pueda generar un
protón.
El efecto túnel mediado por protones ha sido observado en triptamina. Esto sugiere que la
catálisis enzimática podría ser definida más exactamente como una
"barrera", en lugar de como hace el modelo tradicional, donde el
sustrato requiere a la enzima para alcanzar una barrera energética más baja.
Inhibición

{kind=link}
Los inhibidores
competitivos se unen reversiblemente al enzima, evitando la unión del sustrato.
Por otro lado, la unión del sustrato evita la unión del inhibidor. Así pues,
sustrato e inhibidor compiten por la enzima.
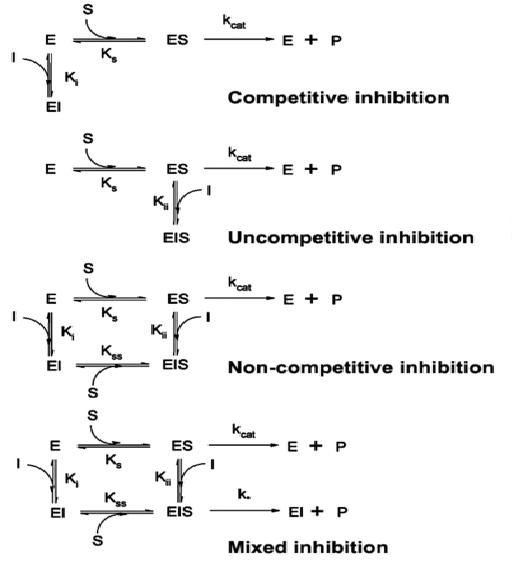
{kind=link}
Tipos de
inhibición según la clasificación introducida por W. W. Cleland.
Los inhibidores
son moléculas que regulan la actividad enzimática, inhibiendo su actividad. A
grandes rasgos, pueden clasificarse en reversibles e irreversibles.
Las irreversibles se unen covalentemente a la enzima sin posibilidad de
revertir la modificación, siendo útiles en farmacología.
Algunos de los fármacos que actúan de este modo son la eflornitina, utilizada
para tratar la tripanosomiasis africana,la penicilina
y la aspirina.
Las reversibles
se unen de forma reversible a la enzima, pudiendo clasificarse a su vez, según
la forma en que intervienen en la reacción, en competitivas, acompetitivas
y mixtas. Habitualmente, por su amplia presencia en multitud de
procesos, se habla también de inhibición no competitiva, que en realidad
no es más que una variante de la ya mencionada inhibición mixta. Sin embargo,
por sus características se suele presentar como opuesta a la competitiva, con
la que es comparada frecuentemente.
·
En la inhibición competitiva, el sustrato y
el inhibidor no se pueden unir a la misma enzima al mismo tiempo, como se
muestra en la figura de la derecha. Esto generalmente
ocurre cuando el inhibidor tiene afinidad por el sitio activo
de una enzima en el cual también se une el sustrato; el sustrato y el inhibidor
compiten para el acceso al sitio activo de la enzima. Por ejemplo, el metotrexato
es un inhibidor competitivo de la enzima dihidrofolato reductasa, que cataliza la
reducción de dihidrofolato a tetrahidrofolato. La similitud entre las
estructuras del ácido fólico y el metotrexato permite que se establezca
una inhibición de tipo competitivo. Este tipo de inhibición se puede superar
con concentraciones suficientemente altas del sustrato, es decir, dejando fuera
de competición al inhibidor. En la inhibición competitiva la velocidad máxima
de la reacción no varía, pero se necesitan concentraciones más elevadas de
sustrato para alcanzar una determinada velocidad, incrementándose así la Km
aparente.
En la inhibición acompetitiva
el inhibidor no puede unirse a la enzima libre, sino únicamente al complejo
enzima-sustrato (ES). Una vez formado el complejo con el inhibidor (EIS) la
enzima queda inactiva. Este tipo de inhibición es poco común, pero puede darse
en enzimas multiméticas.
La inhibición no competitiva es una forma
de inhibición mixta donde la unión del inhibidor con la enzima reduce su actividad pero no afecta la unión con el
sustrato. Como resultado, el grado de inhibición depende solamente de la
concentración de inhibidor, independientemente de la concentración de sustrato,
con lo que varía el valor de la Vmax aparente. Sin embargo, como el
sustrato aún puede unirse a la enzima, el valor de Km no varía.
·
En la inhibición mixta, el inhibidor se puede
unir a la enzima al mismo tiempo que el sustrato. Sin embargo, la unión del
inhibidor afecta la unión del sustrato, y viceversa. Este tipo de inhibición se
puede reducir, pero no superar al aumentar las concentraciones del sustrato.
Aunque es posible que los inhibidores de tipo mixto se unan en el sitio activo,
este tipo de inhibición resulta generalmente de un efecto alostérico donde el inhibidor se
une a otro sitio que no es el sitio activo de la enzima. La unión del inhibidor
con el sitio alostérico cambia la conformación (es decir, la estructura terciaria) de
la enzima de modo que la afinidad del sustrato por el sitio activo se reduce.
La coenzima ácido fólico
(izquierda) y el fármaco anti-cancerígeno metotrexato
(derecha) son muy similares en estructura. Como resultado, el metotrexato es un
inhibidor competitivo de muchas enzimas que utilizan folato.
En muchos
organismos, los inhibidores pueden actuar como parte de un mecanismo de
realimentación. Si una enzima produce una sustancia en demasiada cantidad en el
organismo, esta misma sustancia podría actuar como un inhibidor de la enzima al
inicio de la ruta que lo produce, deteniendo así dicha producción cuando haya
una cantidad suficiente de la sustancia en cuestión. Este sería una forma de realimentación negativa. Las enzimas que
se encuentran sujetas a este tipo de regulación suelen ser multiméricas y
poseer sitios alostéricos donde se unen sustancias reguladoras. Las gráficas que
representan la velocidad de la reacción frente a la concentración de sustrato
de estas enzimas no son hipérboles, sino sigmoidales (forma de S).
Usos de los
inhibidores
Debido a que
los inhibidores modulan la función de las enzimas, suelen ser utilizados como
fármacos. Un típico ejemplo de un inhibidor que es utilizado como fármaco es la
aspirina,
la cual inhibe las enzimas COX-1 y COX-2 implicadas en la síntesis de un intermediario
inflamatorio, las prostaglandinas, con lo que suprime así los
efectos derivados, el dolor
y la inflamación.
Sin embargo, otros inhibidores enzimáticos actúan como venenos. Por
ejemplo, el cianuro
es un inhibidor irreversible que se une a los átomos de hierro y cobre en el sitio activo
de la citocromo c oxidasa de células animales (las plantas son
resistentes al cianuro), bloqueando así la respiración celular.
Función biológica
Las enzimas
presentan una amplia variedad de funciones en los organismos vivos. Son
indispensables en la transducción de señales y en procesos de
regulación, normalmente por medio de quinasas
y fosfatasas.También son capaces
de producir movimiento, como es el caso de la miosina
al hidrolizar
ATP para generar la contracción muscular o el movimiento de
vesículas por medio del citoesqueleto.Otro tipo de
ATPasas en la membrana celular son las bombas de iones implicadas
en procesos de transporte activo. Además, las enzimas también
están implicadas en funciones mucho más exóticas, como la producción de luz por la luciferasa en las luciérnagas.Los virus también pueden
contener enzimas implicadas en la infección celular, como es el caso de la integrasa
del virus HIV
y de la transcriptasa inversa, o en la liberación
viral, como la neuraminidasa del virus de la gripe.
Una importante
función de las enzimas es la que presentan en el sistema
digestivo de los animales. Enzimas tales como las amilasas
y las proteasas
son capaces de degradar moléculas grandes (almidón
o proteínas,
respectivamente) en otras más pequeñas, de forma que puedan ser absorbidas en
el intestino.
Las moléculas de almidón, por ejemplo, que son demasiado grandes para ser
absorbidas, son degradadas por diversas enzimas a moléculas más pequeñas como
la maltosa,
y finalmente a glucosa,
la cual sí puede ser absorbida a través de las células del intestino.
Diferentes enzimas digestivas son capaces de degradar diferentes tipos de
alimentos. Los rumiantes
que tienen una dieta herbívora, poseen en sus intestinos una serie de
microorganismos que producen otra enzima, la celulasa,
capaz de degradar la celulosa presente en la pared celular
de las plantas.
Varias enzimas
pueden actuar conjuntamente en un orden específico, creando así una ruta
metabólica. En una ruta metabólica, una enzima toma como sustrato el
producto de otra enzima. Tras la reacción catalítica, el producto se transfiere
a la siguiente enzima y así sucesivamente. En ocasiones, existe más de una
enzima capaz de catalizar la misma reacción en paralelo, lo que permite
establecer una regulación más sofisticada: por ejemplo, en el caso en que una
enzima presenta una actividad constitutiva pero con una baja constante de
actividad y una segunda enzima cuya actividad es inducible, pero presenta una
mayor constante de actividad.
Las enzimas
determinan los pasos que siguen estas rutas metabólicas. Sin las enzimas, el
metabolismo no se produciría a través de los mismos pasos, ni sería lo
suficientemente rápido para atender las necesidades de la célula. De hecho, una
ruta metabólica como la glucolisis no podría existir sin enzimas. La glucosa,
por ejemplo, puede reaccionar directamente con el ATP de forma que quede
fosforilada en uno o más carbonos. En ausencia de enzimas, esta reacción se
produciría tan lentamente que sería insignificante. Sin embargo, si se añade la
enzima hexoquinasa
que fosforila el carbono 6 de la glucosa y se mide la concentración de la
mezcla en un breve espacio de tiempo se podrá encontrar únicamente glucosa-6-fosfato
a niveles significativos. Por tanto, las redes de rutas metabólicas dentro de
la célula dependen del conjunto de enzimas funcionales que presenten.
Control de la actividad
La actividad
enzimática puede ser controlada en la célula principalmente de estas cinco
formas:
Producción de la enzima (a nivel de la transcripción o la traducción): la síntesis de una enzima
puede ser favorecida o desfavorecida en respuesta a determinados estímulos
recibidos por la célula. Esta forma de regulación génica se denomina inducción
e inhibición enzimática. Por ejemplo, las bacterias
podrían adquirir resistencia a antibióticos como la penicilina
gracias a la inducción de unas enzimas llamadas beta-lactamasas,
que hidrolizan el anillo beta-lactámico de
la molécula de penicilina. Otro ejemplo, son las enzimas presentes en el hígado
denominadas citocromo P450 oxidasas, las cuales son de
vital importancia en el metabolismo de drogas y fármacos.
La inducción o inhibición de estas enzimas puede dar lugar a la aparición de interacciones farmacológicas.
Compartimentalización de la enzima: las enzimas pueden localizarse en
diferentes compartimentos celulares, de modo que puedan tener lugar diferentes
rutas metabólicas de forma independiente. Por ejemplo, los ácidos grasos
son sintetizados por un conjunto de enzimas localizadas en el citosol,
en el retículo endoplasmático y en el aparato de
Golgi, y posteriormente, dichos ácidos grasos son utilizados por
otro conjunto de enzimas diferentes como fuente energética en la mitocondria,
a través de la β-oxidación.
·
Inhibidores y activadores enzimáticos: las enzimas pueden ser activadas o
inhibidas por ciertas moléculas. Por ejemplo, el producto final de una ruta
metabólica suele actuar como inhibidor de alguna de las enzimas implicadas en
las primeras reacciones de la ruta, estableciendo así una realimentación negativa que regula la
cantidad de producto final obtenido por esa ruta. Este mecanismo de
realimentación negativa permite ajustar efectivamente la velocidad de síntesis
de los metabolitos intermedios con la demanda de la célula, y permite
distribuir económicamente materiales y energía para evitar exceso o escasez de
los productos finales. Este control enzimático permite mantener un ambiente
relativamente estable en el interior de los organismos vivos.
·
Modificación postraduccional de enzimas: las enzimas pueden sufrir diversas
modificaciones postraduccionales como la fosforilación,
la miristoilación y la glicosilación.
Por ejemplo, en la respuesta a insulina, se produce la fosforilación de multitud de enzimas,
como la de la glucógeno sintasa, que
ayuda en el control de la síntesis o degradación del glucógeno
y permite a la célula responder a las variaciones de los niveles de azúcar
en sangre.Otro ejemplo de
modificación postraduccional es la degradación de la cadena polipeptídica. La quimiotripsina,
una proteasa
digestiva, es sintetizada en una forma inactiva, quimiotripsinógeno, en el páncreas
y transportada en este estado hasta el estómago,
donde será activada. De este modo se evita que la enzima digiera el páncreas y
los demás tejidos por los que pasa antes de llegar al estómago. Este tipo de
precursor inactivo de una enzima es denominado zimógeno.
Activación dependiente del ambiente: algunas enzimas pueden ser activadas
cuando pasan de un ambiente con unas condiciones a otro con condiciones
diferentes, como puede ser el paso del ambiente reductor del citoplasma
al ambiente oxidativo del periplasma, el paso de un ambiente con elevado pH a otro con bajo pH,
etc. Por ejemplo, la hemaglutinina del virus de la gripe es activada mediante
un cambio conformacional que se produce cuando el pH del medio es
suficientemente ácido, lo cual ocurre cuando el virus entra en el interior de
la célula a través de un lisosoma.
Implicaciones en enfermedades
Estructura tridimensional
de la enzima fenilalanina hidroxilasa (PDB
1KW0).
Debido a que es
necesario un fuerte control de la actividad enzimática para la homeostasis,
cualquier fallo en el funcionamiento (mutación, incremento o reducción de la
expresión o deleción) de una única enzima crítica puede conducir al desarrollo
de una enfermedad genética. La importancia de las
enzimas se pone de manifiesto en el hecho de que una enfermedad letal puede ser
causada por el mal funcionamiento de un único tipo de enzima de todos los miles
de tipos que existen en nuestro cuerpo.
Un ejemplo de esto es el tipo más común de fenilcetonuria. En esta enfermedad genética se produce una mutación de un único aminoácido en la fenilalanina hidroxilasa, una enzima que cataliza la primera reacción de la ruta de degradación de la fenilalanina y de compuestos relacionados. Al ser esta enzima inactiva, se acumulan una serie de productos que terminan dando lugar a la aparición de retardo mental si no se recibe tratamiento.
Otro ejemplo es
cuando se produce una mutación en los genes de la línea germinal que codifican
las enzimas implicadas en la reparación del ADN. En este caso, al no
repararse adecuadamente el ADN de las células, se acumulan mutaciones que
suelen derivar en el desarrollo de diversos tipos de cáncer hereditarios, como
la xerodermia pigmentosa.
Clasificación y nomenclatura de enzimas
El nombre de
una enzima suele derivarse del sustrato o de la reacción química que cataliza,
con la palabra terminada en -asa. Por ejemplo, lactasa
proviene de su sustrato lactosa; alcohol deshidrogenasa proviene de la
reacción que cataliza que consiste en "deshidrogenar" el alcohol;
ADN
polimerasa proviene también de la reacción que cataliza que consiste
en polimerizar el ADN.
La Unión
Internacional de Bioquímica y Biología Molecular ha desarrollado una
nomenclatura
para identificar a las enzimas basada en los denominados Números EC.
De este modo, cada enzima queda registrada por una secuencia de cuatro números
precedidos por las letras "EC". El primer número clasifica a la
enzima en base a su mecanismo de acción. A continuación se indican las seis
grandes clases de enzimas existentes en la actualidad:
·
EC1 Oxidorreductasas: catalizan reacciones de
oxidorreducción o redox.
Precisan la colaboración de las coenzimas de oxidorreducción (NAD+, NADP+, FAD) que aceptan o ceden
los electrones
correspondientes. Tras la acción catalítica, estas coenzimas quedan modificadas
en su grado de oxidación, por lo que deben ser recicladas antes de volver a
efectuar una nueva reacción catalítica. Ejemplos: deshidrogenasas,
peroxidasas.
·
EC2 Transferasas: transfieren grupos
activos (obtenidos de la ruptura de ciertas moléculas) a otras
sustancias receptoras. Suelen actuar en procesos de interconversión de monosacáridos,
aminoácidos,
etc. Ejemplos: transaminasas, quinasas.
·
EC3 Hidrolasas: catalizan reacciones de hidrólisis
con la consiguiente obtención de monómeros
a partir de polímeros. Actúan en la digestión de los alimentos,
previamente a otras fases de su degradación. La palabra hidrólisis se
deriva de hidro → 'agua' y lisis → 'disolución'. Ejemplos:
glucosidasas, lipasas, esterasas.
·
EC4 Liasas: catalizan reacciones en las que se eliminan grupos H2O,
CO2 y NH3 para formar un doble enlace o añadirse a un
doble enlace. Ejemplos: descarboxilasas, liasas.
·
EC5 Isomerasas: actúan sobre determinadas moléculas obteniendo de ellas
sus isómeros
funcionales o de posición, es decir, catalizan la racemización y cambios de
posición de un grupo en determinada molécula obteniendo formas isoméricas.
Suelen actuar en procesos de interconversión. Ejemplo: epimerasas
(mutasa).
EC6 Ligasas:
catalizan la degradación o síntesis de los enlaces denominados
"fuertes" mediante al acoplamiento a moléculas de alto valor
energético como el ATP. Ejemplos: sintetasas, carboxilasas
No hay comentarios:
Publicar un comentario